Tree Dominated Wildlife Habitats
Subalpine Conifer
(SCN) Engelmann Spruce, Subalpine Fir, Mountain Hemlock
Vegetation
Structure-- Typical SCN habitats are open forests with needle-leaved evergreen trees of low to medium stature. Stand density and tree height are typically greater at lower limits of its elevational range (Küchler 1977). In protected sites at lower elevations, tree height may exceed 30 m (100 ft), but trees on exposed sites and windy ridges near tree line are shaped into krummholz stunted, mat-like forms often only about 1 m (3 ft) tall. Shrubby vegetation and herbaceous ground cover are generally sparse or lacking. Litter accumulation is typically scanty, but fallen woody material persists for long periods in the cold climate.
Composition-- Several species dominate canopies of this type in different localities, either singly or in mixtures of two or more species (Parsons 1980). These include Engelmann spruce, subalpine fir, mountain hemlock, western white pine, lodgepole pine, whitebark pine, foxtail pine, bristlecone pine, and limber pine. Although typically of minor importance, a shrub understory may include Parry manzanita, squaw currant, purple mountain heather, oceanspray, and big sagebrush. Willows, western huckleberry, California huckleberry, Sierra bilberry, and alpine laurel occur on moist sites. Western wheatgrass, California brome, several species of lupines, and a variety of flowering annuals are common in the sparse ground cover (Rundel et al. 1977, Parsons 1980). Other Classifications-- SCN combines several types described by other authors. It includes the Engelmann Spruce-Alpine Fir, Mountain Hemlock, Whitebark Pine, Foxtail Pine, Western White Pine, Limber Pine, and Bristlecone Pine of Parker and Matyas (1979) and the Society of American Foresters (SAF) (Eyre 1980), as well as California Mixed Subalpine (256) of the SAF. Subalpine Coniferous Forests (8.6) of Cheatham and Haller (1975) include approximately the same range of dominants. Bristlecone Pine, Foxtail Pine, and Limber Pine of Paysen et al. (1980) are included, as are the Subalpine Forest (17) and Bristlecone Pine Forest (18) or Munz and Keck (1959). Finally Upper Montane-Subalpine Forest (17), Southern Montane-Subalpine Forest (18), and Great Basin Subalpine Forest (19) of Küchler (1977) overlap with SCN as described here.
Habitat Stages
Vegetation Changes-- 1;2-5:S-D. Most high-elevation forests in California have been little disturbed by human influence, but fire, wind, and other natural disturbances offer chances to study successional changes in these forests. Owing to their lack of economic importance, however, few such studies have been carried out in SCN habitat. Parsons (1980:90) observed that the infrequent fire pattern at high elevations in the Sierra Nevada leads to "a specific vegetational mosaic...characteristic of the high country." No details are available. Most of the canopy dominants are pioneers and probably also climax species, at least in harsher environments. For example, whitebark and limber pine, though less shade tolerant than subalpine fir, Engelmann spruce, and mountain hemlock, can regenerate without wildfire or logging due to their longevity and ability to survive extremely cold climates. Furthermore, the short growing seasons, as well as the typically open nature of the stands at higher elevations help ensure regeneration (Pfister et al. 1977).
Duration of Stages-- Little information is available, although Parsons (1980:90) notes that "the slow growth and longevity of most of the subalpine species lead to long time spans between stages." Many tree species that dominate stands of this type are well known for their longevity and slow growth. A whitebark pine 43 cm (17 in) in dbh was 800 years old (Arno 1967), foxtail pine has been aged at nearly 2000 years (Mastroguiseppe 1972), and bristlecone pine has been aged at over 4600 years in the White Mountains of eastern California and over 4900 years on Wheeler Peak in eastern Nevada (Hawksworth and Bailey 1980).
Biological Setting
Habitat-- SCN intergrades with Lodgepole Pine, Jeffrey Pine, and Red Fir habitats at lower elevations. The shrub understory and ground cover are better developed where SCN habitats adjoin moist sites, as along riparian corridors or montane meadows.
Wildlife Considerations-- Coniferous forests at high elevations in California typically support fewer species of amphibians, reptiles, birds, and mammals than any other major forest type in the State. The reasons, though, not clearly established, probably involve some combination of climate, short growing season, lower primary productivity, moisture stress, and lower production of insects and other invertebrates that provide food resources for many vertebrates. Excluding species dependent on ponds, lakes, streams, or cliffs, Laudenslayer (1980) shows no amphibians and only one reptile that find conditions suitable for breeding in these high-elevation forests. He lists only 17 species of birds and 15 of mammals that find conditions optimum, and 14 birds and 22 mammals that find conditions suitable for breeding in such forests. Several species that find optimum or suitable conditions at these high elevations have special significance for land managers. These include the great gray owl, pileated woodpecker, marten, and wolverine.
Burney (1980:99) writes that "birds and small mammals consume so much limber pine seed that little is left for natural regeneration. However, small mammals that transport and bury seed aid dissemination into new areas, and germination of forgotten seed caches undoubtedly accounts for dense groups of stems occasionally found." Such groups of stems are better known to grow from caches left by Clark's nutcrackers, which are robably the primary agents of seed dispersal for limber and perhaps whitebark pines (Tomback and Kramer 1980, Tomback 1982).
Physical Setting
These forests typically occupy extremely harsh environments. Soils are generally thin and of low quality coarse sand, gravel, volcanic debris, and rocks derived from decomposing parent material. Although subalpine fir and Engelmann spruce occur together on rocky, moist moraines and are considered to be indicators of high soil moisture (Parker and Matyas 1979), most stands of SCN are on dry, well-drained soils. The climate is especially challenging. For example, in the Subalpine Forest type described by Munz and Keck (1959), precipitation averages only 76 to 127 cm (30 to 50 in) and may be as low as 38 cm (15 in) east of the crest of the Sierra Nevada. Heavy snow cover is usual. Mean summer high temperatures probably do not exceed 18 C (65 F), and killing frosts are possible during all months. The growing season lasts only 7 to 9 weeks. Similarly, in bristlecone pine forests, measured precipitation over a 3-year period averaged 38 cm (15 in), much of it as snowfall. Mean summer maximum temperatures ranged from 12 to 19 C (54 to 66 F), and winter minima ranged from 16 to 6 C (3 to 21 F) (Munz and Keck 1959). Intense winds are characteristic of these habitats.
Distribution
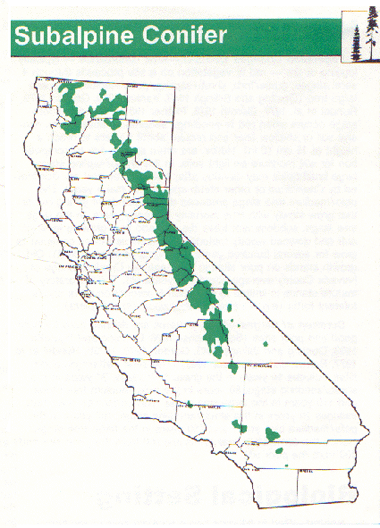
SCN is generally distributed at high elevations in all significant mountain ranges of the State. It is well represented in the north, with an elevational range from about 2,100 to 2,900 m (7,000 to 9,500 ft); and in the Sierra Nevada, ranging from 2,700 to 3,350 m (9,000 to 11,000 ft). "It is poorly represented in southern California" (Parsons 1980:90), typically within an elevational range of 2,880 to 3,400 m (9,500 to 11,200 ft) (Cheatham and Haller 1975). Engelmann spruce-subalpine fir stands are poorly represented in California, occurring together only near Russian Peak in southwestern Siskiyou County (Griffin and Critchfield 1972, Parker and Matyas 1979). "Mountain hemlock, lodgepole pine, western white pine, and California red fir are common in mixed stands above the red fir forest in the north" (Parsons 1980:90). Mountain hemlock is the usual dominant in the northern Sierra Nevada, gradually disappearing to the south (Rundel et al. 1977). The southernmost stand occurs in Tulare County (Parsons 1972). In the Warner Mountains and from about Lake Tahoe southward in the Sierra Nevada, whitebark and lodgepole pine dominate stands of SCN to about the headwaters of the Kern River, where foxtail pine becomes the usual dominant species (Cheatham and Haller 1975). Whitebark pine tends to be the dominant conifer near tree line in the central Sierra Nevada (Cheatham and Haller 1975, Rundel et al. 1977). Limber pine occupies similar sites on the east side of the Sierra Nevada in Inyo and Mono Counties (Arno 1967, Griffin and Critchfield 1972), and together with lodgepole pine it comprises the dominant type in stands of SCN in southern California (Cheatham and Haller 1975). Bristlecone pine stands occur from about 2900 to 3500 m (9500 to 11,500 ft) elevation in the White and Inyo Mountains and the Last Chance and Panamint Ranges, east of the southern Sierra Nevada (Parker and Matyas 1979).
Literature Cited
Arno, S. F. 1967. Interpreting the timberline. M.S. thesis. Univ. of Montana, Missoula. Cheatham, N. H., and J. R. Haller. 1975. An annotated list of California habitat types.
Univ. of California Natural Land and Water Reserve System, unpubl. manuscript Eyre, F. H., ed. 1980. Forest cover types of the Unites States and Canada. Soc. Amer. Foresters, Washington D.C.
Griffin, J. R., and W. B. Critchfield. 1972. The distribution of forest trees in California. U.S. Dep. Agric., For. Serv. (Berkeley, Calif), Res. Pap. PSW-82.
Hawks, F. G., and D. K. Bailey. 1980. Bristlecone pine. 209 Pages 89-90 In F. H. Eyre, ed. Forest cover types of the United States and Canada. Soc. Amer. Foresters, Washington, D.C.
Kuchler, A. W. 1977. Appendix: the map of the natural vegetation of California. Pages 909-938 In M. G. Barbour and J. Major, eds, Terrestrial vegetation of California. John Wiley and Sons, New York.
Mastroguiseppe, R. J. 1972. Geographic variation in foxtail pine Pinus balfouriana. Grev. & Balf. M.S. Thesis. Humboldt State Univ., Arcata, Calif. Munz, P. A., and D. D. Keck. 1959. A California flora. Univ of California Press, Berkeley.
Parker, I., and W. J. Matyas. 1979. CALVEG: A classification of Californian vegetation. U.S. Dep. Agric., For. Serv., Reg. Ecol. Group. San Francisco.
Parsons, D. J. 1972. The southern extensions of Tsuga mertensiana (mountain hemlock) in the Sierra Nevada. Madrońo 21:536-539.
Parsons, D. J. 1980. California Mixed Subalpine 256. Pages 90-91 In F. H. Eyre, ed. Forest cover types of the United States and Canada. Soc. Amer. Foresters, Washington, D.C.
Paysen, T. E., J. A. Derby, H. Black, Jr., V. C. Bleich, and J. W. Mincks. 1980. A vegetation classification system applied to southern California.U.S. Dep. Agric., For.Serv., (Berkeley, Calif.) Gen. Tech. Rep. PSW-45.
Pfister, R. D., B. L. Kovalchik, S. F. Arno, and R. C. Presby. 1977. Forest habitat types of Montana. U.S. Dep. Agric., For. Serv., (Ogden, Utah), Gen. Tech. Rep. INT- 34.
Rundel, P. W., D. T. Gordon, and D. J. Parsons. 1977. Montane and subalpine vegetation of the Sierra Nevada and Cascade Ranges. Pages 559-599 In M. G. Barbour and J. Major, eds. Terrestrial vegetation of California. John Wiley and Sons, New York.
Tomback, D. F.1982. Dispersal of whitebark pine seeds by Clark's nutcrackers: a mutualism hypothesis. J. Anim. Ecol. 51:451-467.
Tomback, D. F., and K. A. Kramer. 1980. Limber pine seed harvest by Clark's nutcracker in the Sierra Nevada: timing and foraging behavior. Condor 82:467- 468.
California Wildlife Habitat Relationships System California Department of Fish and Game California Interagency Wildlife Task Group Subalpine Conifer Jared Verner and Kathryn L. Purcell
Grand Canyon Wildlife
... the Mohave desert scrub regions along the Colorado River in the park’s western end to the Kaibab Plateau’s subalpine conifer forests on the North Rim. ...
Bighorn Sheep
... sagebrush, bitterbrush, pinyon-juniper, palm oasis, desert riparian, desert succulent shrub, subalpine conifer, perennial grassland, montane chaparral, ...
Bats - Death Valley Wildlife
Found in upper creosote bush desert to subalpine confifer forest. ... Most often found in conifer forests; rapid, direct flier; hunts at tree tops. ...
Water in Yosemite National Park
Above 9600 feet, alpine and subalpine zones have little vegetation and low soil .... and upland terrestrial ecosystems, such as conifer forests. ...
Vegetation of Yosemite National Park
The upper montane forest is replaced by the subalpine forest near 8000 feet (2450 m), .... and upland terrestrial ecosystems, such as conifer forests. ...
San Jacinto Mountains: Southern California Mountains and Valleys
... Mixed conifer series on the southwest side of the San Jacinto Mountains, ... Mixed subalpine forest series, Mixed conifer series, Parry pinyon series, ...
Upper San Gabriel Mountains: Southern California Mountains
... Mixed conifer series on the south side, and Jeffrey pine series on the north ... Lodgepole pine series, Mixed conifer series, Mixed subalpine forest ...
Sierra Nevada Ecological Subsections
Predominant potential natural communities include the Mixed conifer series, ... Aspen series, Big sagebrush series, Mixed subalpine forest series, ...
Upper San Gorgonio Mountains: Southern California Mountains
... Lodgepole pine series, Mixed conifer series, Mixed subalpine forest series, Mountain juniper series, Singleleaf pinyon series, White fir series. ...