Shrub Dominated Wildlife Habitats
Montane Chaparral
(MCP) Ceanothus Species, Manzanita Species, Bitter Cherry
Vegetation
Structure-- The growth form of montane chaparral species can vary from treelike (up to 3 meters) to prostrate. When mature, it is often impenetrable to large mammals. Its structure is affected by site quality, history of disturbance (e.g., fire, erosion, logging) and the influence of browsing animals. For example, on shallow granitic soils in the Sierra Nevada, low dense growths of pinemat manzanita and huckleberry oak characterize an edaphic climax community, associated with scattered conifers and much exposed granite. Following fire in the mixed conifer forest habitat type, whitethorn ceanothus-dominated chaparral may persist as a subclimax community for many years. Montane chaparral is characterized by evergreen species; however, deciduous or partially deciduous species may also be present. Understory vegetation in the mature chaparral is largely absent. Conifer and oak trees may occur in sparse stands or as scattered individuals within the chaparral type.
Composition-- Montane chaparral varies markedly throughout California. Species composition changes with elevational and geographical range, soil type, and aspect. One or more of the following species usually characterize montane chaparral communities: whitethorn ceanothus, snowbrush ceanothus, greenleaf manzanita, pinemat manzanita, hoary manzanita, bitter cherry, huckleberry oak, sierra chinquapin, juneberry, fremont silktassel, Greene goldenweed, mountain mahogany, toyon, sumac and California buckthorn. As one or more of these species become dominant under various environmental regimes, further subclassification of the montane chaparral series is possible (Krebs 1972, McNaughton 1968).
Other Classifications-- Montane chaparral has been broadly described as chaparral (Munz and Keck 1973, (Küchler 1977) or mountain shrub (USDA 1977). Subclassifications based upon predominant species composition have also been described as montane mixed shrub series, huckleberry oak/pinemat manzanita series, bush chinquapin series, greenleaf manzanita series, tobacco brush series, mountain whitethorn series (Parker and Matyas 1981); upper montane chaparral, lower montane chaparral (Cheatham and Haller 1975).
Habitat Stages
Vegetation Changes-- 1;2-4:S-D. Montane chaparral in California occurs in gradations between two characteristic successional sequences: The first sequence is associated with poorer, typically shallow soils (in early stages of development), often overlying fractured bedrock. Here, chaparral species may predominate to form an edaphic climax community.
In the second sequence, chaparral is a secondary succession following disturbance on deeper forest soils. After disturbance (logging, fire, erosion) chaparral proliferates and may exclude conifers and other vegetation for many years. However, chaparral may facilitate the germination of red fir seedlings (Barbour 1984) and other shade tolerant conifers by providing a protective cover, moderating microclimate, and improving soil conditions. Chaparral shrubs may be an essential link in forest succession by building up soil nutrient levels, especially nitrogen, to the point where trees can survive (Zavitovski and Newton 1968). In mature timber stands, chaparral species may senesce due to insufficient light through the canopy and are only present as a sparse understory. Thus, silvicultural practices have a strong influence on the structure of montane chaparral. Most montane chaparral species are fire adapted. Mature plants sprout back from the root crown. Some species require scarification of the seed for germination and may produce numerous seedlings after a fire (Gratkowski 1961). However, if fires are too frequent, these species may be eliminated (Biswell 1969) changing the subsequent structure of the community. Deer and livestock foraging on sprouting chaparral may also have a significant effect on its rate of development, structure, and ultimate species composition (Biswell and Gilman 1961, Davis 1967). The forage yields of most sprouting shrubs are reduced for the first few years after a fire, but rapidly regain their original status. Burned areas commonly produce new shrub growth high in protein and are a preferred food source for herbivores (Einarsen 1946, Swank 1956).
Duration of Stages-- Following fire, herbaceous plants may dominate for up to 5 years. Usually within 7 to 9 years the brush overstory is fully developed (Sweeney 1956, Sampson 1944). Chaparral may persist for up to 50 years or longer before conifer development begins to significantly reduce the shrub growth through shading (Lyon 1969, Sweeney 1968). Where chaparral types occur as an edaphic climax (i.e., on poor, rocky soils, fractured bedrock or lava caps), growth rates may be rather slow, growth form is usually small and stunted, and individuals may be quite old. Development of montane chaparral at high elevations is often slowed by cold temperatures, snow cover and a short growing season (Barbour and Major 1977). However, at lower elevations, burned or logged areas may sprout new growth by the next growing season.
Biological Setting
Habitat-- Montane chaparral adjoins a variety of other wildlife habitats, including montane riparian (MRI), mixed chaparral (MCH), and perennial grassland (PGS). It becomes established in disturbed coniferous habits such as ponderosa pine (PPN), mixed conifer (SMC), Jeffrey pine (JPN), red fir (RFR) and lodgepole pine (LPN). At high elevations in the southern Sierra, it may occur with a sparse juniper overstory. At the lower extent of its elevational range, montane chaparral may intergrade with mixed chaparral, a very similar habitat type.
Wildlife Considerations-- Montane chaparral provides habitat for a wide variety of wildlife. Numerous rodents inhabit chaparral (Wirtz 1974). Deer and other herbivores often make extensive use of chaparral. Throughout the west slope of the Sierra and south through the Transverse Range, deer are strongly associated with chaparral communities. Montane chaparral provides critical summer range foraging areas, escape cover and fawning habitat. In the Sierra, fawning areas are frequently found where the chaparral lies adjacent to or contains an interspersion of perennial grass or meadow-riparian habitat (Ashcraft 1975, Dasmann, 1971, Ashcraft 1976, Pacific Gas and Electric 1981). Some small herbivores use chaparral species in fall and winter when grasses are not in abundance. Rabbits and hares eat twigs, evergreen leaves and bark from chaparral. Shrubs are important to many mammals as shade during hot weather, and moderate temperature and wind velocity in the winter (Loveless 1967). Many birds find a variety of habitat needs in the montane chaparral. It provides seeds, fruits, insects, protection from predators and climate, as well as singing, roosting and nesting sites (Verner and Boss 1980), Storer and Usinger 1970).
Physical Setting
Montane chaparral can be found on shallow to deep soils, on all exposures, and from gentle to relatively steep slopes. It may dominate on more xeric sites, but occurs locally throughout the coniferous forest zone. Generally, climate is like that associated with the coniferous forest zone, cold winter temperatures with substantial precipitation. Summers are typically hot and dry (Barbour and Major 1977). In the northern portion of the state, montane chaparral is found between 914 to 2743 m (3000-9000 ft). In southern California this type occurs above 2134 m (7000 ft).
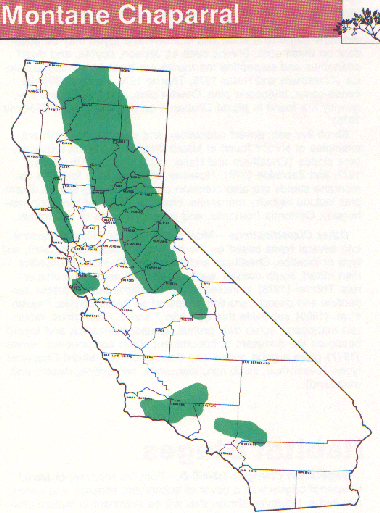
Distribution
Montane chaparral is associated with mountainous terrain from mid to high elevation at 914 to 3047 m (3000-10,000 ft). It occurs in southern California above 2134 m (7000 ft) in the Transverse Range of Los Angeles, and in San Bernardino, Riverside and San Diego counties; from Siskiyou to Kern counties in the Cascade and Sierra Nevada mountains; as a minor type from Tehama to Lake counties; and in Del Norte, Siskiyou, Trinity, and Shasta counties in the North Coast Ranges and Klamath mountains (Barbour and Major 1977). As a successional stage following disturbance, its distribution coincides with the ponderosa pine and mixed coniferous forest habitat types (Barbour and Major 1977).
Literature Cited
Ashcraft, G. C. 1975. Wildlife population response to habitat manipulation: comparative deer use in patchcuts and virgin forest types. California Dep. Fish and Game (Sacramento), Job Prog. W-51-R-20.
Ashcraft, G., Jr. 1976. Deer propagation units and population centers. California Dep. Fish and Game Sacramento.
Barbour, M. G. 1984. Can a red fir forest be restored? Fremontia 11(4):18-19. Barbour, M. G., and J. Major eds. 1977. Terrestrial vegetation of California. John Wiley and Sons, New York.
Biswell, H. H. 1969. Prescribed burning for wildlife in California brushlands. Trans. North Amer. Wildl. and Natur. Res. Conf. 34:438-446.
Biswell, H. H., and J. H. Gilman. 1961. Brush management in relation to fire and other environmental factors on the Tehama deer winter range. Calif. Fish and Game 47:357-389.
Cheatham, N. H., and J. R. Haller. 1975. An annotated list of California habitat types. Univ. of California Natural Land and Water Reserve System, unpubl. manuscript
Dasmann, W. 1971. Deer habitat management in forests. Pages 91-101 In If deer are to survive. Wildl. Manage. Inst., Washington, D.C.
Davis, J. 1967. Some effects of deer browsing on chamise sprouts after fire. Amer. Midl. Nat. 77:234-238.
Einarsen, A. S. 1946. Crude protein determination of deer food as an applied management technique. Trans. North Amer. Wildl. Conf. 11:309-312.
Gratkowski, H. 1961. Brush seedlings after controlled burning of brushlands in southwestern Oregon. J. Forestry. 59:885-888.
Krebs, C. J. 1972. Ecology: The experimental analysis of distribution and abundance. Harper and Row, New York
Kuchler, A. W. 1977. Appendix: the map of the natural vegetation of California. Pages 909-938 In M. G. Barbour and J. Major, eds, Terrestrial vegetation of California. John Wiley and Sons, New York.
Loveless, C. M. 1967. Ecological characteristics of mule deer winter range. Colorado Dep. Game, Fish and Parks Tech. Publ. 20.
Lyon, L. J. 1969. Wildlife habitat research and fire in the northern Rockies. Pages 213- 227 In Proc. Tall Timbers Fire Ecol. Conf.
McNaughton, S. J. 1968. Structure and function in California grasslands. Ecology 49:962-972.
Munz, P. A., and D. D. Keck. 1973. A California flora with supplement. Univ. of California Press, Berkeley.
Pacific Gas and Electric Co. 1981. Exhibit S. Pages 18-23 In Application for amendment of application for new license, FERC Project 137, Mokelumne River. Pacific Gas and Electric Co., San Ramon, Calif.
Parker, I., and W. J. Matyas. 1981. CALVEG: a classification of Californian vegetation. U.S. Dep. Agric., For. Serv., Reg. Ecol. Group, San Francisco.
Sampson, A. W. 1944. Plant succession on burned chaparral lands in northern California. Univ. Calif. Agr. Exp. Sta. Bull. 685.
Storer, T. l., and R. L. Usinger.1963. Sierra Nevada natural history . . . an illustrated handbook. Univ. of California Press, Berkeley.
Swank, W. G. 1956. Protein and phosphorous content of browse plants as an influence on southwestern deer herd levels. Trans. North Amer. Wildl. Conf. 21:141-158.
Sweeney, J. R. 1956. Responses of vegetation to fire: a study of the herbaceous vegetation following chaparral fires. Univ. Calif. Publ. Bot. 28:143-250.
Sweeney, J. R. 1968. Ecology of some "fire type" vegetation in northern California. Pages 111-125 In Proc. Tall Timbers Fire Ecology Conf.
U.S. Department of Agriculture 1977. Regional timber types: compartment inventory and analysis handbook. U.S. Dep. Agric., For. Serv., Pacific Southwest Region, San Francisco.
Verner, J., and A. S. Boss tech. coords. 1980. California wildlife and their habitats: western Sierra Nevada. U.S. Dep. Agric. For. Serv. (Berkeley, Calif.), Gen. Tech. Rep. PSW-37.
Zavitovski, J., and M. Newton.1968. Ecological importance of snowbrush, Ceanothus velutinus in the Oregon Cascades. Ecology 49:1135-1145.
California Wildlife Habitat Relationships System, California Department of Fish and Game, California Interagency Wildlife Task Group, Montane Chaparral - Roland J. Risser and Michael E. Fry