Shrub Dominated Wildlife Habitats
Chamise-Redshank Chaparral
(CRC) Chamise, Redshank, Ceanothus Species
Vegetation
Structure-- Fire occurs regularly in Chamise-Redshank Chaparral and influences habitat structure. Mature Chamise-Redshank Chaparral is single layered, generally lacking well-developed herbaceous ground cover and overstory trees. Shrub canopies frequently overlap, producing a nearly impenetrable canopy of interwoven branches. Chamise-dominated stands average 1 to 2 m (3.3 to 6.6 ft) in height, but can reach 3 m (9.8 ft) (Horton 1960, Cheatham and Haller 1975, Hanes 1977). Total shrub cover frequently exceeds 80 percent, but may be considerably lower on extremely xeric sites with poor soils (Minnich 1976, Vogl 1976, Hanes 1977). Redshank stands are slightly taller, averaging 2 to 4 m (6.6 to 13.1 ft) but occasionally reaching 6 m (19.7 ft) (Hanes 1965, 1977, Cheatham and Haller 1975). Mature redshank frequently is more open than chamise and can have sparse herbaceous cover between shrubs (Hanes 1965, 1977, Paysen et al. 1980).
Composition-- Chamise-Redshank Chaparral may consist of nearly pure stands of chamise or redshank, a mixture of both, or with other shrubs. The purest stands of chamise occur on xeric, south-facing slopes (Hanes 1976). Toyon, sugar sumac, poison oak, redberry, and California buckthorn are commonly found in drainage channels and on other relatively mesic sites (Vogl 1976). At upper elevations or on more mesic exposures, chamise mixes with ceanothus, manzanita, scrub oak, and laurel sumac (Horton 1960, Hanes 1976, Parker and Matyas 1981). Ceanothus and sugar sumac are common associates of redshank (Hanes 1965, 1977). In southern California, white sage, black sage, and California buckwheat are common at lower elevations and on recently disturbed sites (Hanes 1965, 1977). Distinguishing Chamise-Redshank Chaparral from Mixed Chaparral (MCH) and Coastal Scrub CSC) is a subjective interpretation based on percent cover by chamise and redshank and time since last burn. Paysen et al. (1980) classify chaparral as chamise or redshank if either species is "dominant". Hanes (1977) considers a stand to be chamise if it comprises 50 to 100 percent of total cover and redshank if it comprises 20 to 50 percent of total cover. For purposes of this description and the WHR model (Salwasser and Laudenslayer 1982), a more complex definition is needed which reflects changes in species composition that occur during post-fire recovery and aging. A stand of brush is classified as Chamise-Redshank Chaparral, as opposed to Mixed Chaparral, if any of the following criteria are fulfilled.
1. Any stand with greater than 60 percent relative shrub cover by chamise and redshank.
2. Young stands recovering from fire with greater than 20 percent absolute shrub cover by chamise and redshank, and greater than 75 percent relative shrub cover by these species and relatively short-lived subshrubs such as yerba santa.
3. Any stand with at least 50 percent relative shrub cover by chamise and redshank and greater than 75 percent relative shrub cover by these species and shrubs of intermediate life span such as several species of ceanothus.
Other Classifications-- Most plant ecologists treat stands dominated by chamise and redshank as distinct types (Cheatham and Haller 1975, Thorne 1976, Hanes 1977, Paysen et al. 1980, Parker and Matyas 1981). Horton (1960) further divides chamise into "pure chamise" and "chamise-ceanothus" to reflect the frequent occurrence of mixtures of these shrubs. The Californian mixed chaparral of Cheatham and Haller (1975) includes many stands of Chamise-Redshank Chaparral that also support a significant component of ceanothus and other shrubs.
Habitat Stages
Vegetation Changes-- 1;24:S-D. Fire is the primary disturbance initiating secondary succession in Chamise-Redshank Chaparral. Annuals, perennial herbs, and subshrubs are abundant for several years after a fire. Shrubs begin to appear either as seedlings or rootcrown sprouts beginning the first growing season after burning (Hanes 1971). As the habitat matures, shrub cover and height increase and herbaceous cover declines (Hanes 1971). Relatively short-lived shrubs and subshrubs, such as California buckwheat, common deerweed, and most species of ceanothus, may be absent or rare in older stands (Horton and Kraebel 1955, Hanes 1977). After each fire, populations of these species and post-fire herbs regenerate quickly from the seed bank in the soil (Sweeney 1956). In old unburned stands, species diversity is low, growth rates are slow, long-lived shrubs accumulate dead material, and some shrubs may die (Hanes 1971, Rundel and Parsons 1979).
Duration of Stages-- The general schedule of post-fire recovery in chaparral is described by Menke and Villase�or (1977) and Zedler (1977). Herbaceous cover is dominant for 1 to 3 years. Long- and short-lived shrubs increase in height and cover but canopies generally do not overlap for 3 to 15 years after fire. From 10 to 30+ years, shortlived shrubs die, shrub cover increases, the canopy closes, and dead material begins to accumulate. Rundel and Parsons (1979) found that, in the Sierra Nevada, chamise growth rates declined and accumulation of dead material began after 16 years. Time to senescence is dependent on local site characteristics. In southern California, Hanes (1977) considers chamise older than 60 years to be senescent, but this may occur in 20 to 25 years in northern California (Sampson 1944). Horton (1960) states that pure chamise in the San Bernardino Mountains reaches 25 percent cover in 10 years, 50 percent in 40 years, and 70 percent in 55 years. However, recovery rates and peak cover vary with soil type, climatic regime, and slope. For example, most mesic sites supporting chamise and ceanothus reach 50 percent cover in 10 years and 90 percent cover in 25 years. Some sites may reach 90percent cover in 10 years (T. E. Paysen, pers. comm.). At 50 years, shrub cover in mixed stands of chamise and ceanothus may decline to 80 percent total shrub cover as ceanothus dies (Hanes 1977).
Biological Setting
Habitat-- Chamise-Redshank Chaparral generally occurs below and grades into Mixed Chaparral (MCH). On some sites, Chamise-Redshank Chaparral may form an ecotone with Ponderosa Pine (PPN), Coastal Oak Woodland (COW), or mixed conifer types. In northern California, the lower boundary is with Annual Grassland (AGS) and Blue Oak-Digger Pine (BOP). In southern California, Coastal Scrub (CSC) may form a broad mosaic with Chamise-Redshank Chaparral. Location of the boundary can depend on fire frequency (Hanes 1971). On desert exposures, redshank stands may occur above either Mixed Chaparral (MPC) or Desert Succulent Scrub (DSC) and either above or below Pinyon-Juniper (PJN).
Wildlife Considerations-- Wildlife species found in this habitat type also are found in either Mixed Chaparral (MCH), Montane Chaparral (MCP), Coastal Scrub (CSC) or Sagebrush (SGB) and in shrubs beneath several woodland and forest types. The primary land management consideration is selection of alternative fire management treatments. Long-term fire suppression can lead to stand senescence (Vogl 1977) and declines in deer (Biswell et al. 1952), small mammals (Quinn 1979), birds (Wirtz 1979), and reptiles (Simovich 1979). Most animal populations reach peak densities in the first two or three decades, frequently 1 to 15 years, after a fire. Repeated fires at short intervals could favor crown-sprouting shrubs over obligate seed sprouters (Vogl 1977). Either management extreme could have long-term impacts on wildlife through changes in nutrient availability, soil quality or vegetation composition, structure, and recovery time. Prescribed burning can be an effective management tool, but the effects vary with season of burn (Rundel 1982). Post-fire herbs may be important in immobilizing nitrogen within the chaparral system (Rundel and Parsons 1980). Protecting these herbs from grazing may be important for effective long-term habitat maintenance (Rundel 1982). Populations of most small vertebrates decline sharply or are eliminated when chaparral is converted to grassland (Lillywhite 1977). Active and passive chaparral management programs must tailor management prescriptions to specific site characteristics and project goals.
Physical Setting
Chamise-dominated stands are most common on south- and west-facing slopes; redshank is found on all aspects (Hanes 1965, 1977, Cheatham and Haller 1975). Soils usually are thin with little accumulation of organic material (Cheatham and Haller 1975). Chamise may be a dominant shrub on some serpentine sites (Parker and Matyas 1981). Chamise-Redshank Chaparral is found in a mediterranean climate; rainfall is 38 to 63 cm (15 to 25 in), less than 20 percent of total precipitation falls in summer, and winters are mild (Oruduff 1974). The predominant land forms are steep slopes and ridges (Thorne 1976).
Distribution
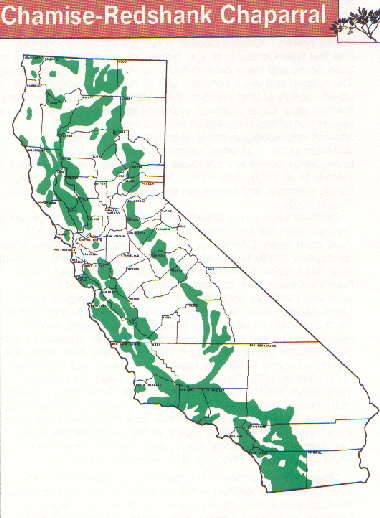
Hanes (1977) provides a good description of "chamise" and "redshank" chaparral distributions in California. This habitat is usually found below 1200 m (4000 ft) on mountain ranges outside the deserts (Cheatham and Haller 1975, Vogl 1976, Minnich 1976, Hanes 1977, Parker and Matyas 1981). Large nearly pure areas of redshankdominated chaparral occur in the interior valleys of the peninsular mountain ranges of Riverside and San Diego counties; isolated stands are found in the Santa Monica Mountains and in northern Santa Barbara and San Luis Obispo counties (Cheatham and Haller 1975, Hanes 1977).Chamise is the dominant shrub of this habitat type throughout the rest of the state. Nearly mature stands of chamise cover large areas in the peninsular and transverse ranges and Tehachapi Mountains of southern California. To the north, chamise more frequently mixes with other shrubs, especially several species of ceanothus. This type of vegetation covers large areas in the central coast ranges and on the eastern exposures of the north coast ranges; as isolated stands in the Cascade and Klamath ranges and the Siskiyou Mountains; and in a broken band on the western slope of the Sierra Nevada (Hanes 1977, Parker and Matyas 1981).
Literature Cited
Cheatham, N. H., and J. R. Haller. 1975. An annotated list of California habitat types. Univ. of California Natural Land and Water Reserve System, unpubl. manuscript Horton, J. S. 1960. Vegetation types of the San Bernardino Mountains, California. U.S.
Dep. Agric., For. Serv. (Berkeley, Calif.), Tech. Pap. No. 44.
Hanes, T. L. 1965. Ecological studies on two closely related chaparral shrubs in southern California. Ecol. Monogr. 35:213-235.
Hanes, T. L. 1971. Succession after fire in the chaparral of southern California. Ecol. Monogr. 41:27-52.
Hanes, T. L. 1976. Vegetation types of the San Gabriel Mountains. Pages 65-76 In J. Latting, ed. Plant communities of southern California. Calif. Native Plant Soc. Spec. Publ. No. 2.
Hanes, T. L. 1977. California chaparral. Pages 417-469 In M. G. Barbour and J. Major, eds. Terrestrial vegetation of California. John Wiley and Sons, New York.
Horton, J. S., and C. J. Kraebel. 1955. Development of vegetation after fire in the chamise chaparral of southern California. Ecology 36:244-262. Lillywhite, H. B. 1977. Effects of chaparral conversion on small vertebrates in southern California chaparral. Biol. Cons. 11:171-184.
Menke, J. W., and R. Villasenor. 1977. The California Mediterranean ecosystem and its management. Pages 257-270 In H. A. Mooney and C. E. Conrad, tech. coords. Symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems. U.S. Dep. Agric., For. Serv., Gen. Tech. Rep. W0-3.
Minnich, R. A. 1976. Vegetation of the San Bernardino Mountains. Pages 99-124 In J. Latting, ed. Plant communities of southern California. Calif. Native Plant Soc. Spec. Publ. No. 2.
Ornduff, R. 1974. Introduction to California plant life. Univ. Of California Press, Berkeley.
Parker, I., and W. J. Matyas. 1981. CALVEG: a classification of Californian vegetation. U.S. Dep. Agric., For. Serv., Reg. Ecol. Group, San Francisco.
Paysen, T. E., J. A. Derby, H. Black, Jr., V. C. Bleich, and J. W. Mincks. 1980. A vegetation classification system applied to southern California. U.S. Dep. Agric., For. Serv., (Berkeley, Calif.) Gen. Tech. Rep. PSW-45.
Quinn, R. D. 1979. Effects of fire on small mammals in the chaparral. Cal-Neva Wild]. Trans. 1979:125-133.
Rundel, P. W. 1982. Successional dynamics of chamise chaparral: the interface of basic research and management. Pages 86-90 In C. E. Conrad and W. C. Oechel, tech. cords. Dynamics and management of Mediterranean-type ecosystems. U.S. Dep. Agric., For. Serv. (Berkeley, Calif.) Gen. Tech. Rep. PSW-58.
Rundel, P. W., and D. J. Parsons. 1979. Structural changes in chamise (Adenostoma fasciculatum) along a fire-induced age gradient. J. Range Manage. 32:462-466.
Rundel, P. W., and D. J. Parsons. 1980. Nutrient changes in two chaparral shrubs along a fire-induced age gradient. Amer. J. Bot. 67:51-58.
Sampson, A. W. 1944. Plant succession on burned chaparral lands in northern California. Univ. Calif. Agr. Exp. Sta. Bull. 685.
Simovich, M. 1979. Post-fire reptile succession. ,Cal-Neva Wildl. Trans. 1979:104-113.
Sweeney, J. R. 1956. Responses of vegetation to fire: a study of the herbaceous vegetation following chaparral fires. Univ. Calif. Publ. Bot. 28:143-250.
Thorne, R F. 1976. The vascular plant communities of California. Pages 1-31 In J. Latting, ed. Plant communities of southern California. Calif. Native Plant Soc. Spec. Publ. 2.
Vogl, R. J. 1976. An introduction to the plant communities of the Santa Ana and San Jacinto Mountains. Pages 77-98 In J. Latting, ed. Plant communities of southern California. Calif. Native Plant Soc. Spec. Publ. No. 2.
Vogl, R. J. 1977. Fire frequency and site degradation. Pages 193-201 In H. A. Mooney and C. E. Conrad, tech. coords. Symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems. U.S. Dep. Agric., For. Serv. Gen. Tech. Rep. W0-3.
Wirtz, W. O., II. 1979. Effects of fire on birds in chaparral. Cal-Neva Wildl. Trans. 1979:114-124.
Zedler, P. H. 1977. Life history attributes of plants and the fire cycle: a case study in chaparral dominated by Cypressus forbesii. Pages 451-458 In H. A. Mooney and C. E. Conrad, tech. coords. Symposium on the environmental consequences of fire and fuel management in Mediterranean ecosystems. U.S. Dep. Agric., For. Serv. (Washington, D.C.) Gen. Tech. Rep. W0- 3.
California Wildlife Habitat Relationships System, California Department of Fish and Game, California Interagency Wildlife Task Group, Chamise-Redshank - Chaparral A. Sidney England